Le Dr Bruno Donatini parle régulièrement des mycorhizes et de l’endobiote dans ses conférences.
Merci à lui car c’est juste passionnant et probablement même un des nutriments clés, dans l’avenir, d’une agriculture performante sans engrais, et de la décontamination des sols pollués.
Mais au fait depuis combien de temps ils existent les champignons ?
Sans les champignons, les arbres n’existeraient pas. La vascularisation des tissus végétaux, nécessaire à l’élévation des plantes, entre autres via la lignine (cette fibre dure qui constitue les troncs), n’existerait pas. Le rendement d’absorption des nutriments par les racines ne serait pas assez performant.
Il a fallu que la nature invente une astuce pour « déléguer » des activations biochimiques : accélérer la fabrication, la synthèse de certains nutriments, notamment l’azote, par une autre espèce vivante.
Il y a environ 450 millions d’années, les champignons apparaissaient : ils allaient révolutionner le règne végétal en permettant un apport nutritionnel supplémentaire déterminant, sur le lieu même d’alimentation de la plante : la racine. A partir de cette époque les plantes ont commencé à se multiplier, et également à grandir, à s’élever dans les airs, créant ainsi une nouvelle compétition : celle de la recherche de la lumière ; les arbres et leurs magnifiques canopées étaient nés. D’abord les fougères géants, les palmiers géants, puis les conifères (gymnospermes). Bien après apparaîtront les plantes à fleur (angiospermes), avec toute la complexité et la magie des interactions avec les insectes.
Ainsi tout le règne vivant allait être impacté : les champignons racinaires (mycorhizes) en sont un des chaînons méconnus, sous-jacents et discrets, puisque souterrains, mais tout à fait essentiels, de l’évolution des plantes.
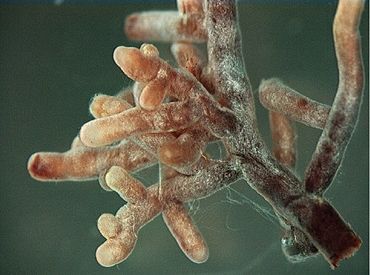
Les mycorhizes. En 1912 dans un mur de pierre sèche au nord de la Grande-Bretagne un médecin écossais remarqua la présence de curieuses roches renfermant des petites plantes et des animaux primitifs. Quelques mois plus tard, un collectionneur de fossiles découvrit qu’elles provenaient d’un village proche : Rhynie (à une quarantaine de kilomètres au nord-ouest d’Aberdeen). Sous la houlette de deux savants britanniques (Robert Kidston et Henry Lang) les fouilles commencèrent aussitôt et débouchèrent sur une découverte sensationnelle : la présence des filaments fongiques d’un champignon archaïque ayant colonisé ces petites plantes primitives.
Cela nous renseigne sur l’écosystème présent à la surface terrestre il y a approximativement 410 millions d’années. Dès cette époque des algues et des champignons entretenaient déjà des interactions complexes relevant de la symbiose mycorhizienne.
Qui est à l’origine de ces derniers termes et que signifient-ils ? Quels sont les effets des mécanismes mis en jeu dans les différents types de symbiose ? Quelles applications en sylviculture, horticulture et maraîchage, agriculture intensive, architecture-paysagiste et myciculture ?
A- Au dix-neuvième siècle, à la suite des nombreux travaux de botanistes européens, naquit l’idée d’une alliance originale entre des champignons et des plantes. Mais c’est à un botaniste-mycologue prussien, Albert Bernard Frank (1839-1900), que revint le mérite d’en faire la synthèse. Il créa le terme mycorhize par l’union de deux mots grecs mukês = champignon et rhiza = racine. Ses motivations étaient d’abord patriotiques. En effet, il travaillait principalement sur des pins sylvestres afin de reproduire leur ensemencement par des spores de truffe noire du Périgord, dans le but de libérer financièrement son pays, la Prusse, des coûteuses importations de ce luxueux produit français. A l’époque la France en produisait plus de 2000 tonnes par an !
Il échoua avec les truffes, mais réussit pleinement avec ses semis de pins cultivés en pot : ses observations furent capitales pour prouver que l’association de champignons sur la racine des plantes (des feuillus et des résineux) relevait non pas d’un phénomène de parasitisme mais d’une association intime (l’un des deux organismes pénètre dans les tissus de l’autre, voire même dans ses cellules), durable et bénéfique aux deux partis concernés. Il forgea alors le terme de symbiose (à partir de deux racines grecques : sym (qui signifie ensemble) et bio (signifiant vie) et formula une théorie révolutionnaire pour son époque : un mycorhize est le siège d’échanges réciproques, une symbiose mutualiste, entre une plante et un champignon. Cette symbiose mycorhizienne nous allons l’étudier de plus près maintenant.
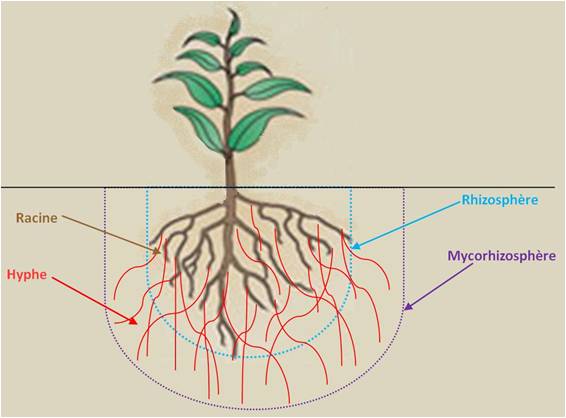
B- De nos jours l’immense majorité des plantes actuelles est associée à des champignons vivant au contact de leurs racines. Par quels mécanismes ? Pour répondre à cette question nous croiserons deux regards : celui du naturaliste et celui du botaniste. Par ce biais nous aborderons ainsi le fonctionnement ses mycorhizes.
D’une façon générale on peut dire ceci : par une mycorhize une plante et un champignon se rendent mutuellement service. A la différence du parasitisme la plante et le champignon sont dans une relation « gagnant-gagnant ». Le mycorhize est constitué à la fois de la racine de la plante et du mycélium du champignon qui enveloppe la racine et la pénètre.
Dans le détail : le champignon étant dépourvu de photosynthèse est incapable de synthétiser du carbone assimilable, la plante va lui en donner d’importantes quantité en échange de nutriments. Et plus précisément encore, le champignon reçoit le carbone au niveau de le mycorhize alors qu’il échange l’eau et les éléments nutritifs du sol (des minéraux qu’il va rendre solubles et des matières organiques présentes dans l’humus qu’il va fragmenter) en les rendant disponibles par la plante sous forme d’azote assimilable par la plante. Le mycélium du champignon, grâce à son réseau très étendu de filaments ténus (des hyphes) multiplie au moins par mille la zone explorée par les racines, permettant ainsi au champignon d’aller fort loin du pied de la plante pour décomposer des molécules complexes de la matière organique présente dans l’humus du sol grâce à ses enzymes spécifiques qui vont également solubiliser des éléments nutritifs comme l’azote, le phosphore présents dans les minéraux des roches. Deux éléments qui selon leur disponibilité sont essentiels à la croissance de la plante. Et comme la majorité des champignons mycorhiziens ne sont pas spécifiques d’une plante, de nombreuses plantes peuvent être connectées par un même réseau de mycélium commun. Un réseau qui peut être immense avec près de 200 mètres d’hyphes mycorhizienne dans un seul gramme de sol forestier !

Du point de vue morphologique le mycélium colonise les racines courtes en s’introduisant plus ou moins profondément sous leur épiderme et les enveloppant comme dans un filet qui porte le nom de réseau de Hartig, en mémoire d’un célèbre naturaliste allemand du 19è siècle. Suivant la forme et la profondeur de pénétration des hyphes et leur forme on distingue quatre types de mycorhize.
Type n° 1 : l’ectomycorhize. Dans une minorité de cas le champignon reste à l’extérieur de sa plante hôte, même s’il lui arrive de pénétrer dans ses tissus. Il enveloppe complètement ses racines par un manteau qui reste bien visible à l’œil nu, ce qui facilita grandement la tâche des chercheurs, dont celle du découvreur de la symbiose ectomycorhizienne : le prussien A.B. Frank déjà cité. De ce manteau sortent des hyphes ou cordons mycéliens vers l’extérieur de la plante assurant les échanges au niveau du sol et vers l’intérieur des hyphes assurant les échanges entre la plante et le champignon. Mais cette symbiose ne concerne qu’une toute petite partie de végétaux (environ 5%) qui occupent cependant de grandes surfaces. Ce sont pour l’essentiel des arbres et des arbustes appartenant à un très petit nombre de familles botaniques visibles aussi bien en milieu forestier dans la zone tempérée de l’hémisphère nord, en zone boréale également (Canada, Scandinavie et Sibérie) où se rencontrent des essences à ectomycorhizes appartenant aux familles suivantes : les Pinacées (ifs, pins, sapins, mélèzes, épicéas), les Bétulacées (aulnes, bouleaux, charmes, châtaigniers et noisetiers), les Fagacées (chênes, hêtres et châtaigniers), les Salicacées (saules, peupliers) et les Tiliacées (tilleuls). Pour la zone méditerranéenne, voire tropicale, ce sont principalement les Cistacées (cistes), les Mimosacées (acacias australiens) et les Myrtacées (des arbres comme les eucalyptus, ou des arbustes tels les mélaleucas endémiques aux zones tropicales du nord de l’Australie, de la Tasmanie, Malaisie, Nouvelle Calédonie et dans le sud-est asiatique.
Leur système racinaire suit un schéma hiérarchisé : d’abord des racines longues qui vont se ramifier en racines courtes. C’est sur ces dernières que le champignon va s’installer et former la symbiose mycorhizienne. Quels champignons ?
La plupart des champignons mycorhiziens sont des Basidiomycètes. On estime à plus de 6000 les espèces de champignons mycorhizeurs. Quelques Ascomycètes sont mycorhiziens, en particulier le genre Tuber avec les truffes. Tous forment des sporophores complexes et de grande taille. Leur diversité s’oppose au petit nombre d’espèces de plantes portant des ectomycorhizes.
Type n° 2 : l’ectendomycorhize. Certains jeunes résineux (les semis de pins, épicéas, mélèzes et de Douglas) sont porteurs d’un autre type de mycorhizes où les hyphes du réseau de Hartig (le champignon est toujours un Ascomycète très spécialisé) sont à la fois à l’extérieur et à l’intérieur des cellules de la racine qu’ils colonisent en perforant la membrane des cellules jusqu’à les occuper entièrement. Mais au bout d’une année cette mycorhize finit par disparaître totalement.
Type n° 3 : l’endomycorhize à pelotons. Son originalité est de former une symbiose avec des plantes poussant sur des sols très pauvres en azote. De quelles plantes s’agit-ils ? Elles sont de deux types :
- d’une part des arbustes de la famille des Éricacées : les bruyères, les callunes, les rhododendrons, les myrtilles, les canneberges, les camarines noires (qui poussent exclusivement en Australie et en Nouvelle-Zélande), les arbousiers et les pyroles qui forment leur symbiose avec de nombreux Basidiomycètes comestibles (des tricholomes, laccaires, lactaires), parfois toxiques (des hébélomes, rhizopogons, ) parfois non comestibles (des téléphores, pisolithes et porias entre autres).
- d’autre part toutes les orchidées. Elles ont les deux particularités suivantes : avoir des graines à la fois minuscules (ce qui favorise la dissémination de l’espèce) et ne pouvoir germer sans l’aide d’un champignon spécifique, car leurs graines sont dépourvues de réserves. Pour les orchidées à feuilles vertes, ce champignon mycorhizien (un Basidiomycète) va leur fournir tous les éléments vitaux en allant décomposer une source de nutriments proches afin d’apporter du carbone sous forme de sucres. Il peut même le prélever directement dans les racines d’une autre plante voisine. Une exception : l’épipactis, orchidée fréquente en forêt européenne, forme ses symbiotes avec des Ascomycètes. Quant aux orchidées non chlorophylliennes, ce qui se remarque à la couleur jaunâtre de leurs feuilles, elles forment des mycorhizes non seulement avec les champignons basidiomycètes précédents, mais également avec des champignons décomposant la matière organique, des sapotrophes, comme les coprins ou les lycoperdons.
Type n°4 : l’endomycorhise arbusculaire : c’est le type de symbiose le plus fréquent (environ 80 % des plantes actuelles : la majorité des arbres fruitiers, les plantes herbacées, les frênes, les érables, etc. et tous les arbres tropicaux ) et c’est également le type de symbiose le plus ancien (car observé sur les algues fossilisées de Rhynie). Cette mycorhize se caractérise par trois aspects : d’un côté l’absence de manteau fongique autour des radicelles, de l’autre par la pénétration des hyphes du champignon partenaire (de la famille des Gloméromycètes) à travers la paroi cellulosique de l’hôte et prennent une forme particulière de petit arbre. D’où leur nom. Enfin, et c’est la dernière caractéristique de cette mycorhize : par l’absence de réseau de Hartig. Cela étant, et malgré leur grand âge, les endomycorhizes arbusculaires restent une des plus grandes réussites du vivant.
Un exemple précis faisant appel à ce type de symbiose se rencontre fréquemment dans les prairies où il a été mis en évidence par l’utilisation de marqueurs isotopiques pour étudier les échanges d’éléments nutritifs entre le trèfle et une graminée. Le premier appartient de la famille des Fabacées comme le pois, le haricot, le soja, etc. et possède la propriété de fixer directement l’azote de l’air grâce aux bactéries symbiotiques de ses racines. La seconde (de la famille des Poacées) en est incapable. Tous deux vont s’interconnecter par le réseau mycélien des champignons Gloméromycètes présents dans le secteur pour effectuer des transferts bidirectionnels de matière (eau, azote, carbone et éléments minéraux).
Cette symbiose compense les inégalités entre espèces. Ainsi est-il courant de rencontrer un même réseau mycélien pouvant couvrir plus de 100 m² et s’associer symbiotiquement avec toutes les plantes de cette surface. Une propriété observée avec tous les végétaux de milieux aussi divers qu’une forêt tempérée, une forêt tropicale humide, ou une prairie.
Il est à noter que certaines plantes n’ont jamais choisi un type de mycorhize particulier. Elles voient la coexistence d’ un type dominant et d’un type minoritaire. Cela se rencontre avec les aulnes, les saules, les peupliers et les eucalyptus dont la symbiose se fait avec une majorité de champignons endomycorhiziens arbusculaires sur les racines longues, tandis que les champignons ectomycorhiziens ne colonisent que les racines courtes des jeunes plantes. Puis la proportion s’inverse et les ectomycorhiziens finissent par coloniser totalement les racines de l’arbre.
Reste une minorité de plantes sans ou avec très peu de mycorhizes. Elle concerne environ 20 % des espèces végétales. Elles vivent soit dans un environnement les dispensant de mycorhize, soit résultent de sélections opérées par l’homme. Trois exemples vont illustrer cette exception.
Viennent en premier des algues totalement immergées comme les posidonies en mer et les élodées en eau douce, ainsi que des plantes dont les racines et les tiges poussent dans l’eau : nénuphars, massettes, roseaux communs ou jacinthes d’eau. Toutes ces plantes sont sans cesse au contact de l’eau salée ou douce et n’ont donc aucun mal à absorber directement cette eau et les nutriments dissous qu’elle renferme.
Le second type de plantes ne présentant pas ou très peu de mycorhizes concerne des variétés de céréales qui sont adaptées aux conditions spéciales de l’agriculture intensive moderne pratiquée dans les pays développés avec une forte fertilisation chimique des sols en NPK (azote, phosphore, potassium).
Arrivent enfin des plantes cultivées comme les choux, les navets, la rhubarbe, le sarrasin, la betterave, l’épinard, le quinoa, l’artichaut, le tournesol et quelques « mauvaises herbes » (le chénopode et la ravenelle). Toutes poussent rapidement, se reproduisent par graines et ont gardé leur faculté de coloniser rapidement des sols érodés ou nus dépourvus d’humus et donc de champignons et de micro-organismes. Elles compensent leur absence de mycorhize en développant un système racinaire soit très vaste, soit très fin, ce qui leur permet d’étendre au maximum leur zone d’échange entre le sol et la plante.
La cohabitation reste cependant possible entre ces différentes plantes : sur une même parcelle lourdement fertilisée artificiellement peuvent se côtoyer du blé, de la betterave et de la ravenelle.
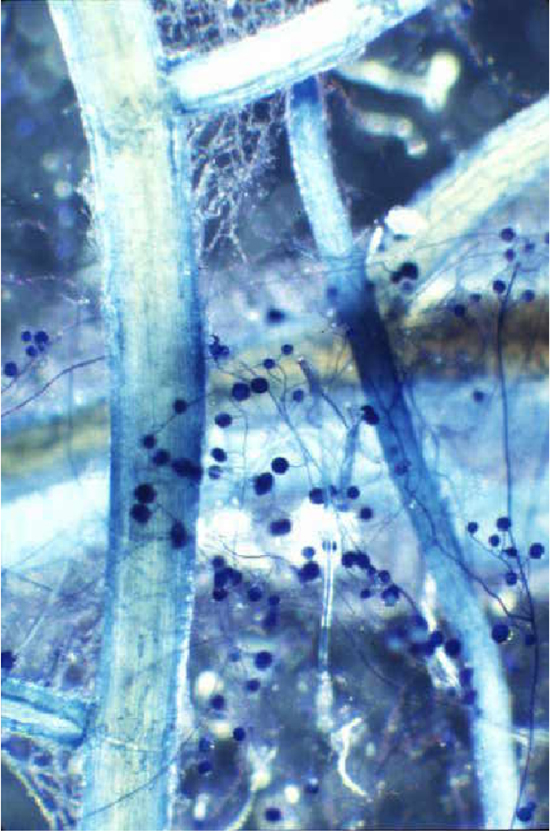
Mais il y a plus : la symbiose mycorhizienne apporte d’autres bénéfices à la plante-hôte. Elle lui procure un système racinaire plus dense, si bien qu’elle pousse plus vite, avec en plus une floraison plus abondante et une fructification prolifique. Elle n’en reste pas là et défend la plante-hôte contre ses adversaires par la gestion de deux types de stress.
D’abord le stress biotique en protégeant ses racines des champignons pathogènes ou même ses tiges et feuilles d’insectes herbivores ! Dans les sols des pépinières forestières vivent aussi bien des champignons favorables à la croissance des semis que des champignons ayant un effet néfaste. Beaucoup sont des espèces appartenant au genre Phytophora, Fuzarium, Pythium, Rhizoctonia pour n’en citer que quelques-uns. Ils deviennent pathogènes quand les semis sont stressés par excès d’arrosage ou par fertilisation excessive, entraînant la fonte des semis ou la nécrose des racines. Pendant longtemps furent employés des désinfectants et des fongicides qui détruisent tous les microorganismes, aussi bien bénéfiques que néfastes. La tendance actuelle consiste à choisir un système de lutte intégrée couplant une utilisation ultra-minimale de pesticides à une lutte biologique faisant appel à des mycorhizes spécifiques comme nous le verrons plus loin en troisième partie.
Voyons à présent comment un mycorhize peut être insecticide. Cette découverte s’est faite fortuitement par des chercheurs néo-zélandais avec le mycélium d’un champignon endophyte (du genre Neotyphodium). Il est présent dans toutes les parties sauf les racines des plantes fourragères consommées par les moutons (diverses fétuques, ray-grass vivace de la famille des Poacées). Par sa production d’un alcaloïde insecticide le mycélium protège doublement ces plantes non seulement des insectes ravageurs détruisant la tige de ces plantes mais aussi de la sécheresse estivale.
Et c’est précisément notre second type de stress annoncé auparavant : le stress abiotique. Il concerne aussi bien la sécheresse estivale, que la sécheresse journalière subie par de nombreuses algues vertes ou brunes de l’estran (bande de terre du littoral tantôt couverte à marée haute, tantôt découverte à marée basse). Dans leur cas le champignon ne pénètre pas les cellules de l’algue, ou de la plante (un eucalyptus par exemple), il la protége par une symbiose ectomycorhizienne à bas potentiel hydrique.
Dernier cas de stress abiotique : le stress résultant de la pollution par hydrocarbures métaux lourds, etc. C’est un autre avantage d’un champignon vivant en symbiose avec son hôte : Pisolithus tinctorius qui est de loin le plus performant car il produit de grandes quantités de complexes chélatés (prononcer Kélaté) se formant à partir d’ions ou de molécules et un atome central de métal. Cette fonction « dépolluante » permet de nettoyer des sols contaminés par des métaux lourds ou de pour restaurer des sols détruits par une activité humaine néfaste aux champignons mycorhizeurs ( terrils de houillères, crassiers de mines de métaux, etc.). Pisolithus tinctorius est le seul à pouvoir encore émettre des symbioses mycorhiziennes avec le pin sur des sols où la température dépasse aisément quarante degrés, comme sur les terrils de houille.
Pour assurer pleinement cette « phytoremédiation » le champion est encore Pisolithus tinctorius qui forme des ectomycorhizes (le mycélium du champignon entoure complètement l’extérieur de la racine et jusqu’à la pénétrer, mais sans franchir les cellules de la plante). Cette symbiose a deux avantages : elle peut explorer de grands volumes de sol soit pour stocker les métaux dans la plante (qu’il sera ensuite possible de recycler pour récupérer les métaux lourds), soit de « casser » les molécules indésirables. Dans ce cas les plantes vont neutraliser les sols pollués. Pour cette tâche les ectomycorhizes sont plus performants que les endomycorhizes : des champignons microscopiques pénétrant alors dans la racine de l’hôte, comme avec les gloméracées trouvées à Rhynie en Écosse,
Quels plantes-hôtes sont employées précisément pour dépolluer des sites industriels ou militaires contaminés et abandonnés ? Selon le climat et le sol de la zone concernée, des pins, des peupliers et des saules ayant recours à des mycorhizes spécifiques.
Abordons maintenant l’effet néfaste d’un mycorhize symbiotique. Une plante le fait fort bien : la callune utilise sa symbiose non pour échanger d’une façon bilatérale avec un champignon spécifique, mais pour éliminer d’autres champignons. Dans toute l’Europe tempérée et boréale elle forme des nappes sur les sols pauvres, acides. Or il se trouve que ces endroits sont souvent choisis par les forestiers pour implanter artificiellement des conifères à croissance rapide comme l’épicéa commun ou le Douglas qu’on peut en voir dans les forêts artificielles d’Écosse, de Bretagne, d’Auvergne ou du Limousin. Pour réussir cette culture les fsylviculteurs ont très vite remarqué qu’il leur fallait impérativement éradiquer la callune en premier par des herbicides ou des travaux mécaniques, faute de quoi leurs jeunes plants restaient jaunes et ne poussaient pas. Mais la cause est plus subtile : la callune forme des mycorhizes spécifiques avec des champignons ascomycètes qui secrètent des substances toxiques non pas pour les racines des jeunes plants, mais pour les champignons ectomycorhiziens spécifiques des épicéas et des Douglas, causant ainsi leur dépérissement. En revanche, cette particularité ne s’applique pas aux pins : ils sont associés à d’autres champignons ectomycorhiziens spécifiques (comme des Rhizopogon) qui restent insensibles aux toxines des symbiotes fongiques de la callune.

C/ Quelques applications de la symbiose mycorhizienne dans 6 domaines :
1/ La sylviculture
Les propriétaires ou les pépiniéristes pépiniéristes désireux d’augmenter leurs productions sans utiliser des engrais coûteux (à l’achat et à la fabrication, côté énergie), voire nocifs pour la nature ont à leur disposition deux voies utilisant la symbiose mycorhizienne : une façon traditionnelle et une façon innovante.
La manière traditionnelle consiste à apporter de la terre ou de l’humus contenant des spores de champignons mycorhizants ; mais ce procédé a l’inconvénient d’être volumineux et surtout risqué par sa possibilité d’introduite des souches pathogènes. Autre possibilité : récolter des spores de Basidiomycètes et d’Ascomycètes, mais cette récolte est délicate et de plus ces spores germent mal. Sauf avec trois groupes de champignons : les truffes, les pisolithes (en climat chaud et avec des sols pauvres) et les rhizopogons qui produisent d’énormes quantités de spores se gardant bien, mais dont la constance génétique n’est pas garantie.
Et de l’autre côté, une façon innovante. En effet, les forestiers plutôt que de continuer à saturer leurs sols d’intrants pour fertiliser des plantes n’utilisant pas ou peu de mycorhizes, ce qui conduit à asphyxier la terre en la privant des ses micro-organismes, ont tout intérêt à mettre en œuvre une MC, c’est à dire une mycorhization contrôlée (traduction directe de l’américain « controlled mycorhization »), concept vulgarisé dans les années 70 en sylviculture par Donald Marx et ses collaborateurs à l’Institut des Recherches sur les Mycorhizes d’Athens en Géorgie. Ils ont sélectionné un champignon aux propriétés remarquables : le Pisolithus tinctorius, champignon acidophile et thermophile qui peut établir des symbioses avec près d’une quarantaine d’autres espèces de champignons. Mais la recherche sur les mycorhizes efficaces est longue, coûteuse, fastidieuse et peu avancée ; mais elle est fiable. Elle a plusieurs applications très prometteuses.
A- le bois à longue rotation avec les trois cas de figure suivants
- en zone tempérée : la majorité des réserves de nutriments est en surface (humus) de sols pauvres en éléments nutritifs facilement assimilables. Si le propriétaire d’une parcelle à boiser ne veut pas dépenser inutilement en engrais chimiques pour amender son sol, il a tout intérêt à utiliser la mycorhization contrôlée effectuée par un pépiniériste spécialisé.
Elle passe par la multiplication in vitro à grande échelle en labo par culture de mycélium pour obtenir des souches pures et abondantes puis par leur inoculation. Cette dernière phase peut avoir lieu sous trois formes : l’inoculant est seul (il faut alors le mélanger à un support de tourbe, terreau) ; l’inoculant est déjà incorporé à un substrat poreux (un mélange de vermiculite expansée et de tourbe) ; enfin l’inoculant est sous forme liquide ou semi-liquide. Il sera apporté lors de l’arrosage des jeunes plants forestiers.
Aux USA vingt-et-une entreprises commercialisent des inoculants endomycorhiziens contre 9 en Europe où la pratique de l’agroécologie est encore balbutiante…mais il est vrai que le retour sur investissement est dissuasif pour un propriétaire puisqu’ entre la plantation et la récolte s’écoulent plusieurs dizaines d’années.
2- en zone tropicale : Il nous faut distinguer deux cas.
Premièrement un environnement humide où sont introduits d’autres espèces d’arbres, comme des résineux. Il faut alors impérativement introduire aussi des champignons compatibles (du genre Pisolithus tinctorius) en mélangeant la terre locale à celle des pépinières d’où proviennent les plants ou mieux encore : utiliser des plants mycorhizés en pépinière. Succès garanti. Autre essence introduite : les eucalyptus.
C’est d’ailleurs la première essence plantée au monde avec des sujets provenant de sélections faites sur des arbres provenant d’Australie, Papouasie ou des îles orientales d’Indonésie. Les records de production au Brésil, en République Populaire du Congo et en Chine méridionale s’expliquent autant par l’amélioration génétique que par la particularité de l’eucalyptus qui forme simultanément ces deux types de symbiose : ectomycorhizienne et endomycorhizienne arbusculaire (cette dernière domine largement en zone tropicale humide). Autre cause du succès de l’acclimatation de l’eucalyptus dans ces pays : collées dans leurs racines les jeunes plants importent des spores de champignons mycorhizeurs originaires d’Australie.
Second cas : un environnement chaud et sec (Sahel, Ouest africain par exemple) sont introduits des acacias (originaires d’Australie) qui fixent l’azote de l’air dans le sol par le biais de leurs mycorhizes, ce qui permet de pratiquer l’agroforesterie avec la cohabitation d’acacias et de mil dans un même champ de culture, ce qui dispense d’engrais comme de fumure organique.
3- En zone tempérée ou boréale : des conditions plus faciles dominent vu que l’essence plantée (un résineux) a toutes les chances de rencontrer de nombreuses symbioses mycorhiziennes compatibles une fois sélectionné les champignons assurant la stabilité génétique de l’inoculant (sous forme de mycélium). Trois exemples vont illustrer la mycorhization contrôlée du sapin de Douglas.
Commençons par la France en notant au passage que c’est la première essence de reboisement dans notre pays au vu de ses grandes qualités (croissance rapide, légèreté, forte résistance mécanique et durabilité : aucun traitement de préservation). Cet arbre bien que privé de ses champignons d’origine, puisqu’il vient de la côte pacifique de l’Amérique du Nord, s’acclimate très bien en Europe en général et en France en particulier où il prospère, principalement dans la région du centre. Les souches les plus performantes sont les Laccaria bicolor avec des résultats fort intéressants pour une mycorhization contrôlée effectuée dans des plantations à une altitude moyenne allant de 300 à 800 m, sur des sols acides (avec un pH inférieur à 5,5) dans des landes à fougères ou après une coupe forestière et non sur d’anciennes terres agricoles. Le mycélium est inoculé soit sous forme solide, soit sous forme liquide. Son efficacité pour améliorer la croissance de cet arbre est moins due à une nutrition minérale améliorée qu’à la fabrication d’auxines favorisant le développement des racines et les protégeant contre les attaques de pathogènes sur ses racines. Pour éviter toute concurrence les pépiniéristes français ne commercialisent pas leur mycélium mais seulement des plants de sapins de Douglas qu’ils ont mycorhizés.
Deuxième exemple : le reboisement de haute montagne dans les Alpes, non pour produire du bois, mais pour fixer le sol et empêcher les éboulements, les glissements de terrain et les avalanches. Témoin la plantation en Autriche de pins cembro mycorhizés par du bolet pleureur.
Troisième et dernier exemple : dans les états du sud-est des USA (la Géorgie, les Carolines du Nord et du Sud, la Floride, etc.) la sylviculture de Douglas mycorhizés par tantôt par Pisolithus tinctorius, sur des sols sablonneux peu fertilisés, tantôt par Hebeloma crustuliniforme qui est plus performant pour des sols plus fertiles.
B/ le bois à courte, voire très courte rotation et le bois d’énergie :
Le but est la production de fagots, rondins de petite taille, plaquettes, pour bois de feu, pâte à papier, biocarburants ou panneaux à particules. Sont concernées toutes les essences forestières capables d’émettre des rejets sur leur souche après une coupe à blanc. Avec une préférence pour les peupliers et les saules en zone tempérée et pour l’eucalyptus en zone tropicale.
Un bémol cependant : il n’existe pas de mycorhization contrôlée pour le bois à très courte rotation.

- L’horticulture et le maraîchage :
Ces deux filières ont plusieurs points communs : des sols riches et une forte valeur ajoutée aux produits, ce qui autorise des coûts de production élevés.
Lorsque c’est possible le sol est désinfecté avant son ensemencement par inoculation. Par quels moyens ? Les trois techniques principales sont les suivantes : utiliser de la vapeur, recourir à des fumigations chimiques avec du bromure de méthyle ou encore chauffer le sol. Avec comme corollaire la destruction totale des champignons résidents, qu’ils soient bénéfiques ou pathogènes, l’élimination des larves d’insectes phytophages vivant dans le sol et la réduction du nombre de vers de terre. En bref, une accumulation d’effets pénalisant la mycorhization de la culture à venir.
S’il est certain que la mycorhization contrôlée n’augmente pas d’une façon spectaculaire la nutrition des plantes cultivées, elle a néanmoins deux avantages : contrôler des champignons pathogènes avec des cultures hors-sol de tomates, poivrons, concombres, etc. sous serre alors que la culture de légumes (carottes, aulx, oignons,etc.) et de fleurs (roses, muguet, œillets, tulipes, iris, etc.) se fait en plein champ. Second avantage : obtenir une croissance optimale par la forte fertilisation des sols.
Il est à noter qu’avec les plantes dites de terre de bruyère (azalées, rhododendrons, bruyères, piéris et myrtilles) la pratique de l’inoculation mycorhizienne est très peu développée.
Par contre, avec les orchidées obtenues par culture de méristème multiplié in vitro, l’association d’une plantule et d’un champignon symbiotique procure un avantage certain.
Terminons par un créneau fort rentable financièrement pour ces deux filières : la diffusion rapide de nouvelles obtentions variétales de fruits et légumes obtenue par l’emploi de la mycorhization contrôlée des très jeunes plants cultivés in vitro. Ainsi sont produits et vendus des clones de petits fruits comme des fraises et des tomates cerises ; sans parler des bananes, de la canne à sucre et des pommes de terre en grande culture.

- L’agriculture intensive
Après la révolution verte qui vit les rendements grimper en flèche grâce aux OGM, aux pesticides, fongicides et désherbants, ainsi qu’à l’emploi d’engrais chimiques déversés à hautes doses (N-P-K), et d’une mécanisation poussée, cette agriculture est dans une impasse : l’érosion s’aggrave par la disparition de la matière organique alimentant la faune et les micro-organismes des sols (le taux de matière organique moyen des sols français a été divisé par deux ou trois selon les régions).
Et les rendements, bien qu’exceptionnels, stagnent, en dépit de nombreuses tentatives d’employer de nouveaux engrais dans les cultures intensives. Si bien que les sols se meurent partout, quel que soit le climat. Ils ne peuvent absorber que 15 % du phosphore apporté et tout le reste disparaît dans le sol !
Or cet élément qui est la clef de voûte de l’ADN et de l’ARN se raréfie de plus en plus dans le monde : dans moins d’un siècle les gisements de phosphates naturels seront épuisés et la demande sera énorme avec l’explosion démographique Il est au centre du métabolisme énergétique de tous les êtres vivants et des plantes en particulier car vital pour la croissance cellulaire et la constitution des graines, du pollen, etc. Comment parer à la crise inévitable ?
Commencer certes par diminuer la consommation de phosphates chimiques en alternant les cultures pour restaurer l’humus et sa vie microbienne est une première étape pour sortir de cette impasse du productivisme agricole où les produits sont certes toujours aussi calorifiques, mais vidés de leurs nutriments essentiels ! Opter pour une agriculture biologique préservant durablement la vie des sols ? Oui, mais à condition de ne pas abuser de fertilisation organique : elle est aussi mauvaise que l’excès d’engrais chimiques pour la rhizosphère en général et la diversité microbienne en particulier.
Mais surtout opter pour solutions agricoles durables en confier l’analyse de ces sols usés à des laboratoires indépendants pour choisir des espèces cultivées selon leur capacité à répondre positivement à la symbiose mycorhizienne. L’utilisation judicieuse de ces mycorhizes permet en effet de solubiliser les phosphates naturels bio-disponibles par la plante. Pratique intéressante pour certains pays pauvres des zones tropicales où les paysans pauvres ne peuvent acheter des engrais chimiques, mais disposent de grands gisements de phosphates naturels.
- L’architecture-paysagiste
Pour les pelouses des particuliers, ou les terrains de golf ou de sport en France, il commence à être possible de se servir des gazons en rouleaux. Ce gazon est composé d’espèces spécifiques de graminées dont les graines sont parfois mycorhizées. Il est vendu au mètre carré en rouleaux de 2,5 m x 0,4. Chaque rouleau pèse une douzaine de kilos pour une épaisseur de 3 cm. Mais avant de le dérouler, il faut commencer par préparer le sol, en le nivelant et en retirant les adventices et les cailloux. Puis, si nécessaire, amender le sol par une couche de terreau mycorhizé. Ensuite dérouler le gazon, le poser en bandes. Après cela le rouler pour le tasser et finir par l’arrosage.
Ce procédé est peu employé en France (moins de 5 % de surfaces engazonnées le sont ainsi) alors que le pourcentage dépasse les 60 % aux USA. Un marché d’avenir ici en France ?
- La myciculture
Ici l’objectif n’est pas de produire plus de plantes, mais plus de champignons comestibles. Le champignon de Paris, le pleurote, le shiitake sont couramment cultivés sur de la paille, du bois ou du compost alors que la truffe, le cèpe et la chanterelle ne peuvent être cultivés sans leur arbre hôte.
Or la sylviculture a un impact direct sur la production de champignons. Il suffira de trois exemples pour le montrer.
- l’épicéa, qui produit beaucoup de cèpes, a souffert de l’ouragan du 26 décembre 1999. Il fut remplacé par le Douglas à l’enracinement plus solide, à la croissance plus rapide et au bois de meilleure qualité. Malheureusement pour les mycologues et les gourmets il n’est pas un hôte favorable à la production de cèpes. : il est rare d’en trouver sous lui.
- la truffe noire du Périgord et le chêne (rouvre, sessile ou pédonculé) : nous l’avons abordé précédemment en relatant les essais infructueux du découvreur de la symbiose ectomycorhizienne en 1885. Il fallut attendre le début des années 1960 pour voir un chercheur italien décrire et réaliser la mycorhize de chênes en trempant leurs racines dans truffes broyées. Ces arbres plantés dans des conditions favorables de sol et d’ensoleillement produisirent des truffes plus vite. Depuis, cette méthode a été étendue à de nombreuses autres espèces de feuillus telles que le noisetier commun, le noisetier de Byzance, le charme, le châtaignier, le tilleul, le bouleau verruqueux. Trois hôtes seulement pour les résineux : le pin noir, le pin sylvestre, et le cèdre de l’Atlas. En France L’INRA commercialise des plants de ces arbres mycorhizés. Récemment sont apparus sur le marché d’autres producteurs de truffes que la France et l’Italie (sa truffe blanche du Piémont atteint des cours astronomiques) avec des pays comme Israël, le Maroc, les USA (la Californie) et la Chine. Ce dernier pays est devenu un réel concurrent pour la truffe française (truffe noire et truffe de Bourgogne), mais ses truffes bon marché sont infiniment moins goûteuses.
- Enfin le pin gris et le matsutake qui, à l’instar de la truffe européenne est le fleuron de la gastronomie japonaise. Mais les tentatives de l’inoculer en pépinière sont restées vaines jusqu’à présent. Il est à signaler que ce champignon très recherché est assez fréquent dans les pinèdes de l’atlas marocain et du nord québécois. Le Canada vient en deuxième place, juste après la Chine, comme pays exportateur de matsutake vers le Japon.
CONCLUSION
Depuis les travaux du botaniste et mycologue prussien Albert Bernhard Frank qui, à la fin du 19è siècle, mit en évidence l’ectomycorhize (une incroyable biotechnologie par laquelle des champignons microscopiques vivant sur les racines de plantes établissent un échange réciproque de nutriments), trois autres symbioses mycorhiziennes au moins ont été découvertes. Dont l’ endomycorhize présente sur plus de 80 % des plantes actuelles. Cette symbiose existait déjà dans des plantes primitives comme l’attestent des fossiles vieux de plus de 400 millions d’années.
La compréhension des interactions bénéfiques que jouent les mycorhizes entre plantes et champignons a permis la réalisation de progrès considérables dans maints domaines allant de la sylviculture, l’agriculture intensive, l’horticulture, le maraîchage, la phytoremédiation, la fabrication de biocarburants et la myciculture pour n’en citer que quelques-uns. Or toutes ces interactions sont une composante fondamentale de la structure et de la fonction de notre écosystème.
La future recherche mycorhizienne pluridisciplinaire devra se poursuivre dans l’approfondissement des mécanismes de ces interactions complexes afin de faire face à six défis : le changement climatique, l’accroissement de la population mondiale, l’impasse de l’agriculture intensive face à des sols souvent appauvris et parfois détruits ainsi que la diminution des ressources mondiales, du phosphate naturel en particulier. Tous les pays sont concernés, qu’ils soient en voie de développement ou plus avancés techniquement. Voilà pourquoi nous sommes tous acteurs de ces futures batailles à livrer. Pour ne pas nous ruiner dans l’achat d’engrais coûteux et peu efficaces, pour ne pas compromettre notre santé avec, pour arrêter de saccager la nature avec des cultures polluantes, nous avons tous intérêt à nous tourner vers des produits et des pratiques saines recourant aux champignons mycorhiziens , solution simple et efficace pour des cultures durables, innovantes et respectueuses de l’individu et de la biosphère. Ce qui nous invite à ne plus appuyer systématiquement sur le champignon, mais à ralentir nos consommations frénétiques de produits douteux et nos cultures intensives polluantes pour vivre mieux, plus longtemps et sauver ainsi notre planète.
Jacques BRALE – Christophe ETIENNE